At the request of - Vivek H. Murthy - 7/15/2021 - Surgeon General of the United States - I want my viewers to know:
Hans and Susan Neuhart (nee CassAdy) were fully vaccinated - for SARS-COV-2 - in May of 2021.
It did not hurt a bit. We had no significant reaction (at all) - and, it does feel good to know [that] we are protected.
We are doing well - and, fully intend to return - to the YMCA - when everyone is vaccinated. SO PLEASE GET VACCINATED !
The White House : Office of the Press Secretary : For Immediate Release
December 02, 2014
Remarks by the President [Barack Obama] on Research for Potential Ebola [Corona Virus Family] Vaccines
National Institutes of Health
Bethesda, Maryland
4:54 P.M. EST
Barack Obama: THE PRESIDENT: Thank you, everybody! It is good to be back. Thank you. Thank you so much. Everybody, please have a seat. Thank you.
Well, to Secretary Burwell, to Francis Collins, Tony Fauci, your teams, to all of you, thanks so much for welcoming me here today. It is wonderful to be back to America’s laboratory, even if I don’t always understand what you’re doing. (Laughter.)
Last year, I welcomed Francis and some of you to the White House to launch our BRAIN Initiative to unlock the mysteries of the mind and to pursue new cures for disease.
And Francis promoted me at the time to “scientist in chief.” (Laughter.) Which made me very proud, although I sort of felt guilty that I hadn’t studied more chemistry. (Laughter.)
But the work you do here is remarkable, and I just got a fascinating tour of your vaccine research center. I have to say, I was very impressed with how you can clone a virus gene into a vaccine vector, then subject it to gel electrophoresis. (Laughter and applause.) And then pipet the samples into a 96-well microplate. (Laughter.) Run it through the world’s most advanced multiparameter flow cytometer. (Laughter and applause.) I mean, it was impressive. (Laughter.) I’ve been tinkering around the White House, setting up a similar system. (Laughter.) We use it for brewing beer. (Laughter.) But it works well for your work also. (Laughter.)
Now, the last time I was here at NIH, early in my presidency, I came to announce a historic boost in funding for biomedical research. Because part of American leadership in the world -- one of the things that has always marked us as exceptional -- is our leadership in science and our leadership in research. And here at NIH, you have always been at the forefront of groundbreaking innovations. You’ve helped pioneer new treatments for everything from cancer to heart disease to HIV/AIDS. And as a consequence, you’ve helped not just Americans but people around the world live longer, fuller lives. You’ve saved countless lives in every corner of the globe. And so to Francis and Tony, and all your directors and staff, and the researchers that you fund across the country and around the globe, you deserve great thanks for your leadership, and your service, and your patriotism, and your lifesaving work.
And that brings me back to today. This past summer, as Ebola spread in West Africa, I told my team that fighting this disease had to be a national security priority, and a priority across agencies and across our government. I realize that here in the United States, some of the attention has shifted away recently -- that’s sort of how our attention spans work sometimes. Ebola is not leading the news right now. But I wanted to come here because, every day, we’re focused on keeping the American people safe. Every day, the NIH is at the forefront of this mission. NIH personnel have volunteered and deployed to West Africa. Some have served in medical labs, testing for Ebola. Some of your clinicians -- members of the U.S. Public Health Service -- have deployed to care for health care workers who got infected in the line of duty.
When Nina Pham, one of the two Dallas nurses who were infected, needed treatment, Tony and his team stepped up and you were ready.
You manned shifts around the clock, day and night. You remembered your training. You displayed great skill and professionalism. You reminded the world that it is possible to treat Ebola patients effectively and safely without endangering yourselves or others.
And all that has made an enormous difference. Like a lot of Americans, I know you fell in love with Nina-- she was so sweet and big smile, her optimism, her sense of service, and reminded us -- she reminded us of the incredible sacrifices that our tireless nurses make every day, and we can never thank them enough. And I know Tony thanks Nina for teaching him how to FaceTime. (Laughter.) And after she was released, Ebola-free, I was proud to welcome Nina to the Oval Office and give her a big hug, and she’s now back home in Texas, recovering, getting stronger. And we remember what she told the world when she was released: “Throughout this ordeal, I have put my trust in God and my medical team.” And we thank everyone on her team at the NIH Clinical Center who delivered such remarkable care to Nina.
But the point is, is that the work that you have done has continued even if the cameras have gone elsewhere. And the urgency remains, because if we are going to actually solve this problem for ourselves, we have to solve it in West Africa as well. And one of the great virtues of what you’ve done here at NIH is reminded people that science matters and that science works. It’s not always going to be immediate; sometimes it’s going to be iterative and there are going to be some trials and there are going to be some errors and false starts and blind alleys, but the basic concept of subjecting hypotheses to tests and seeing if they work and being able to document them and replicate them -- the basic concept of science -- and making judgments on the basis of evidence, that’s what’s most needed during difficult, challenging moments like the ones that we had this summer and that we continue to have in West Africa.
Last week, just in time for Thanksgiving, NIH and your partners gave us something new to be thankful for, and that was news of the first successful step -— completion of the Phase 1 clinical trials -- of a potential Ebola vaccine. And on my tour just now, Doctors Nancy Sullivan and Mario Roederer showed me how they and their teams did it. And I have to say both Nancy and Mario were really good teachers and were very patient with my rudimentary questions, and the lasers were really cool. (Laughter and applause.)
No potential Ebola vaccine has ever made it this far. So this is exciting news. But it’s also a reminder of the importance of government-funded research and our need to keep investing in basic research. (Applause.)
Because Nancy, as she was talking about the steps that had been taken, showed me -- this is the kind of mementos scientists keep I guess -- is there was some numbers on a little chart -- (laughter) -- from back in 1999? -- in which she had first done some experiments and trials on the Ebola virus. So this is the product not just of last year’s work; it’s the product of over a decade of inquiry and work. And at the time, when -- Nancy was explaining when she first had some breakthroughs in understanding the Ebola virus, nobody really gave a hoot. Until you do.
And that’s part of how science works -- you make investments and you pursue knowledge for knowledge’s sake, in part because it turns out that knowledge may turn out useful later and you don’t always know when.
Last week’s news is still just a first step. There are no guarantees. But Dr. Cliff Lane, who is here, is working with Liberian officials to begin large-scale tests in that country. And other potential Ebola vaccines are also in the works. I know that here at NIH you’re also working on potential treatments for Ebola. As you move ahead on all these fronts, I want you to know you have your President’s full support, and the administration’s full support.
You are a vital part of our fight against Ebola, across our government. Today, we released an update on our efforts, here in the United States and abroad. And it shows that, because we’ve stepped up our efforts in recent months, we’re more prepared when it comes to protecting Americans here at home. We’re screening and monitoring arrivals from the affected countries. We’ve equipped more hospitals with new protective gear and protocols. We’ve conducted outreach and training of hundreds of thousands of healthcare workers.
A few months ago, only 13 states could test for Ebola; today 36 states can. Previously, there were only three facilities in the country deemed capable of treating an Ebola patient, including NIH. Today, we’re announcing that we now have 35 Treatment Centers designated to care for a patient with Ebola. So this is important progress. And we’re going to just keep on at it. And throughout, we are going to be guided by the science -- not by speculation, not by fear, not by rumor, not by panic -- by science.
Now, part of what the science and epidemiology and experience has taught us -- and I’ve said this all along -- is the best way to fight this disease, to protect Americans, is to stop it at its source.
And that’s why the United States continues to lead the global response in West Africa. Some 3,000 of our servicemembers and civilians are now on the ground -- manning that air bridge, moving in supplies, building treatment units. I called some of our troops in West Africa on Thanksgiving to express gratitude and they were inspiring, the can-do spirit that they displayed.
The new Medical Unit we built in Liberia to treat health workers opened last month and has begun discharging patients Ebola-free. We’ve ramped up the capacity to train hundreds of new health workers per week. We’ve helped improve burial practices across Liberia. And as a consequence, we’ve seen some encouraging news: A decline in infection rates in Liberia. And meanwhile, over the last few months, the United States has helped rally the international community.
We’ve mobilized more than $2 billion in commitments to this fight because this has to truly be a global effort. But that money would not be there had it not been for U.S. leadership.
So our strategy is beginning to show results. We’re seeing some progress. But the fight is not even close to being over. As long as this disease continues to rage in West Africa, we could continue to see isolated cases here in America. In West Africa, this remains the worst Ebola epidemic in history by a long shot. And although we’ve made some progress in Liberia, we’ve still got work to do. We are seeing that we still have a lot of work in Guinea, and it’s actually been getting worse in Sierra Leone despite some good efforts from our British partners. And this can still spread to other countries, as we’ve seen in Mali. Every hot-spot is an ember that, if not contained, could become a new fire. So we cannot let down our guard, even for a minute. And we can’t just fight this epidemic; we have to extinguish it.
Much of the progress we’ve made -- and the progress we still need to make -- depends on funding that’s running out. We can’t beat Ebola without more funding. This is an expensive enterprise. And that money is running out. We cannot beat Ebola without more funding. If we want other countries to keep stepping up, we will have to continue to lead the way.
And that’s why I’m calling on Congress to approve our emergency funding request to fight this disease before they leave for the holidays. It’s a good Christmas present to the American people and to the world. (Applause.)
The funding we’re asking for is needed to keep strengthening our capacity here at home, so we can respond to any future Ebola cases. The funding allows us to keep making progress in West Africa. Remember, we have to extinguish this disease -- this is not something that we can just manage with a few cases here and there; we’ve got to stamp it out. The funding is needed to speed up testing and approval of any promising Ebola vaccines and treatments, including those here at the NIH. It’s needed to help us partner with other countries to prevent and deal with future outbreaks and threats before they become epidemics.
This is something I want to just focus on for a second. Tony and I were fondly reminiscing about SARS and H1N1. (Laughter.) That’s what these guys do for fun. (Laughter.) And we were lucky with H1N1 -- that it did not prove to be more deadly. We can’t say we’re lucky with Ebola because obviously it’s having a devastating effect in West Africa -- but it is not airborne in its transmission.
There may and likely will come a time in which we have both an airborne disease that is deadly. And in order for us to deal with that effectively, we have to put in place an infrastructure -- not just here at home, but globally -- that allows us to see it quickly, isolate it quickly, respond to it quickly. And it also requires us to continue the same path of basic research that is being done here at NIH that Nancy is a great example of.
So that if and when a new strain of flu, like the Spanish flu, crops up five years from now or a decade from now, we’ve made the investment and we’re further along to be able to catch it. It is a smart investment for us to make. It’s not just insurance; it is knowing that down the road we’re going to continue to have problems like this -- particularly in a globalized world where you move from one side of the world to the other in a day.
So this is important now, but it’s also important for our future and our children’s future and our grandchildren’s future. And the last few elections, the American people have sent Washington a pretty clear message: Find areas where you agree, don’t let the areas where you disagree shut things down, work together and get the job done.
I cannot think of a better example of an area where we should all agree than passing this emergency funding to fight Ebola and to set up some of the public health infrastructure that we need to deal with potential outbreaks in the future. How do you argue with that? That is not a partisan issue. That is a basic, common-sense issue that all Americans can agree on.
Now, I have to say I’ve been very encouraged so far by the bipartisan support in our various visits with members of Congress. For the most part, people have recognized this is not a Democratic issue or a Republican issue -- it’s about the safety and security of the American people. So let’s get it done. This can get caught up in normal politics -- we need to protect the American people and we need to show the world how American leads.
I have to tell you, I traveled to Asia, we had the G20 Summit -- if America had not led, if I had not been able to go to CDC, make a major announcement about the commitments we were going to make, be able to go to the United Nations and basically call on other countries to step up, and know that we were following through with our own commitments, had we not done that, the world would not have responded in the same way. American leadership matters every time. We set the tone and we set the agenda.
Now, in closing, I want to leave with a story that speaks to what we have to do. Nancy Writebol, is from Charlotte, North Carolina. She’s a mom, grandma, wife, also a Christian missionary. Along with her husband, she went to Liberia. She was doing God’s work -— caring for Ebola patients. It’s hard to imagine a greater expression of the Christian ethic. And she was then infected herself. So she was brought back to Emory in Atlanta, she received excellent care. Nancy was released in August. She is Ebola-free, she continues to recover. And she said this about how people treat her, even today: “You have some people that just totally wrap their arms around you, and shake your hand. And then you have other people that stand 10 feet away.”
Some people wrap their arms around you. Some people stand 10 feet away. This disease is not just a test of our health systems; it is a test of our character as a nation.
It asks us who we are as Americans. When we see a problem in the world -- like thousands of people dying from a disease that we know how to fight -- do we stand 10 feet away, or 10,000 miles away, or do we lead and deploy and go to help?
And I know what kind of character I want to see in America, and I know the kind of character that’s displayed by people here at NIH and some of your colleagues that are deployed right now in Liberia -- that’s who we are. We don’t give in to fears. We are guided by our hopes and we are guided by our reason, and we are guided by our faith, and we’re guided by our confidence that we can ease suffering and make a difference. And we imagine new treatments and cures, and we discover, and we invent, and we innovate, and we test, and we unlock new possibilities.
And when we save a life and we help a person heal, we go up to them and we open our arms, and we wrap our arms around them with understanding and love and compassion and reason. That’s what you do here at NIH. It’s what we do as Americans. That’s who we are. That’s who we’ll always be.
Thank you very much. God bless you. God bless the United States of America. (Applause.)
Gorbalenya, A.E., Krupovic, M., Mushegian, A. et al." The new scope of virus taxonomy: partitioning the virosphere into 15 hierarchical ranks." > Nat Microbiol5, 668–674 (2020). https://doi.org/10.1038/s41564-020-0709-x
Abstract : Virus taxonomy emerged as a discipline in the middle of the twentieth century. Traditionally, classification by virus taxonomists has been focused on the grouping of relatively closely related viruses. However, during the past few years, the International Committee on Taxonomy of Viruses (ICTV) has recognized that the taxonomy it develops can be usefully extended to include the basal evolutionary relationships among distantly related viruses. Consequently, the ICTV has changed its Code to allow a 15-rank classification hierarchy that closely aligns with the Linnaean taxonomic system and may accommodate the entire spectrum of genetic divergence in the virosphere. The current taxonomies of three human pathogens, Ebola virus, severe acute respiratory syndrome coronavirus and herpes simplex virus 1 are used to illustrate the impact of the expanded rank structure. This new rank hierarchy of virus taxonomy will stimulate further research on virus origins and evolution, and vice versa, and could promote crosstalk with the taxonomies of cellular organisms.
Main: Viruses were discovered at the end of the nineteenth century as filterable agents causing infectious diseases of plants and animals1,2,3,4,5. Subsequently, their pathogenicity and ability to undergo rapid evolutionary change6 has sparked a large body of research, often connected to the so-called ‘microevolution’ of relatively closely related viruses7,8. However, over the last decade, our appreciation of the importance and distribution of viruses has expanded beyond the original parasitic–pathogen model, and now virologists recognize the role of viruses in host regulation and the maintenance of natural ecosystems9. Shotgun metagenomic sequencing has also revealed the presence of a vast variety of viruses in diverse environmental samples and in apparently healthy organisms from all divisions of life10,11,12,13. ( REFERERENCES, Aclnowledgements, contributors, etc )
To understand the true extent of virus genomic diversity—which may be significantly broader than that of their hosts—and the origins and forces that shape this diversity, virologists will have to systematically rationalize the more distant relationships between viruses, ideally reflecting their ‘macroevolution’, and virus taxonomy should provide an inclusive yet dynamic classification framework to reflect these relationships. In contrast to the taxonomies of cellular organisms, this new virus taxonomic framework will have to accommodate the current view that viruses have multiple origins (polyphyly) and that their diversity cannot be represented by a single virosphere-wide tree14.
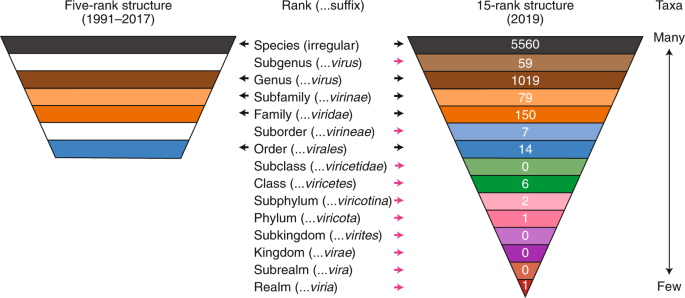
The traditional five-rank structure of virus taxonomy : The International Committee on Taxonomy of Viruses (ICTV) oversees the official classification of viruses and nomenclature of taxa, that is, taxonomy (Box 1)15. In its earliest versions, the ICTV classification of viruses into taxa formally recognized only genera and families but, over time, this classification scheme developed into a five-rank hierarchy of species, genus, subfamily (used rarely), family and order16,17. This five-rank structure matched a section of the Linnaean hierarchical structure used in the taxonomies of cellular organisms and remained in place until 2017 (Fig. 1, left). In addition to changes in the rank hierarchy, the recognition of virus taxa has also evolved over time, from a traditional phenotype-based characterization process to a multistage process that increasingly, but not exclusively, includes genomic properties and sequences (Box 2)15,18. Nowadays, formal virus classification emphasizes comparative sequence analyses of conserved genes and proteins, including gene phylogeny, gene synteny and shared gene content. Other molecular traits are also considered when appropriate19,20.
Fig. 1: A comparison of the ICTV taxonomic rank hierarchy in 1991–2017 and 2019.
Taxonomic ranks are shown in relation to the distribution pattern of taxa. The number of taxa assigned to each rank (as recorded in the current ICTV Master Species List, release 2018b, MSL34 (ref. 47)) are shown in white font on the 15-rank structure. When the ranks are described as a hierarchy, the species rank is often referred to as the lowest rank and the realm rank as the highest rank. However, when the ranks are used as phylogenetic terms, the realm rank can be described as basal and the species rank as apical or terminal. Both conventions are used in this Consensus Statement. Black arrows, ranks common to the five- and 15-rank structure; pink arrows, ranks introduced in the 15-rank structure.
Classifications outside of the ICTV taxonomic remit : Until recently, the evolutionary relationships between viruses of different families or orders were considered by the ICTV, and by many in the virology community, as being too distant to be resolved in a credible classification. Thus, there was little impetus to extend the taxonomy rank structure. The result was a taxonomy that, in profound contrast to its cellular counterparts, included many disjointed taxa, the number of which increases with the accelerating discovery of novel viruses (exceeding 100 families in 2018). However, classification efforts continued outside of the official taxonomic framework and, over the last few decades, several informal groupings such as ‘supergroups’ or ‘superfamilies’ were proposed for subsets of RNA viruses21,22,23 and DNA viruses24,25,26,27,28. These groupings relate to otherwise seemingly disparate viruses belonging to different families and have a variety of different hosts, genome types and organizations, and replication mechanisms. Importantly, these groupings have relied on distant relationships often associated with structure–function hypotheses (for example, an essential virus protein involved in virus replication or virion morphogenesis), which were then validated in subsequent experimental studies29,30,31; these provided independent support for the inferred classifications. ( REFERERENCES, Aclnowledgements, contributors, etc )
Also, before these developments took place, Baltimore had introduced a non-hierarchical classification of viruses which groups viruses into just seven (originally six) classes according to their genome type (double-stranded DNA, single-stranded DNA, double-stranded RNA, positive-sense RNA, negative-sense RNA, reverse-transcribing RNA and reverse-transcribing DNA) and its relation to the synthesis of mRNA32,33. Because of its conceptual clarity and functional foundation, this classification system is still widely used. It complements virus taxonomy by grouping viruses into meaningful classes at a different scale of virus divergence, albeit without attempting to evaluate their evolutionary relationships.
The new Linnaean-like ranking hierarchy of virus taxonomy : In 2016, the urgency, timeliness and logistics of introducing additional ranks to the virus taxonomy hierarchy were discussed at length by the ICTV Executive Committee (EC). The discussion addressed how best to mirror the complete Linnaean taxonomy system (based on a nest of seven principal or primary ranks: species, genus, family, order, class, phylum and kingdom), how to allow for the hierarchical clustering of virus taxa in higher ranks such as orders, and whether the Baltimore classes might be adopted as taxa, perhaps at the basal ranks of the taxonomy34. Figuratively speaking, a taxonomic hierarchy was sought that could accommodate a virosphere-wide tree (or trees) from the roots to the tips of the branches. Because of its potential impact on the practice of virus taxonomy, the EC created a Working Group to consider the matter in more detail. An account of the process undertaken by the Working Group to propose a new taxonomy is outlined in Box 3.
The Working Group concluded that an extended, formal virus classification hierarchy should provide 15 ranks, including eight principal (or primary) ranks and seven derivative (or secondary) ranks (Fig. 1). The eight principal ranks include four that were already in use (order, family, genus and species) and four that are new: realm, kingdom, phylum and class, which are all above the order rank. The class rank in this series is not to be confused with the ‘classes’ described by Baltimore, or the typological attributes of a taxonomic rank35. These new principal ranks cover the entire scale of virus divergence to include the deepest virus relationships at the basal rank of realm. The large scale of virus divergence encompassed by the 15 ranks is exemplified by the newly created Riboviria taxon (a realm) that currently includes all RNA viruses encoding an RNA-directed RNA polymerase, including viruses of three Baltimore classes (III, IV and V)36.
The seven secondary ranks include the previously used subfamily rank and six new ranks that are derivatives of most of the remaining principal ranks. The exception is the species rank, which is currently not associated with a secondary rank, as no consensus on the definition of ‘subspecies’ could be reached. This new rank hierarchy and the associated nomenclature (Fig. 1, right), including defined suffixes for taxa, follow those used in the Linnaean system with a single exception. The basal rank is called ‘realm’ in virus taxonomy, rather than ‘domain’ (as in other taxonomies), reflecting a complex interrelation between virus taxonomy and its counterparts for cellular organisms.
The new rank hierarchy and its normative basis, in the form of changes to the ICTV Code, were approved by the ICTV EC and subsequently ratified by the ICTV in two votes in 2018 and 2019 (refs. 37,38). These changes provide the virology community with the opportunity to submit taxonomic proposals that fill the new principal and secondary ranks with defined taxa.
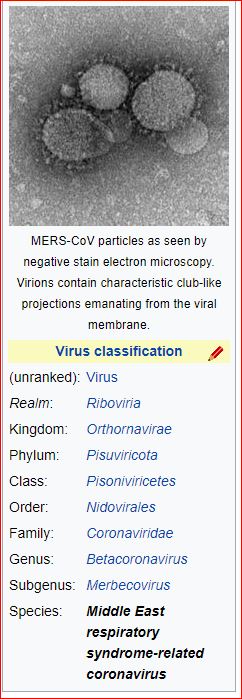
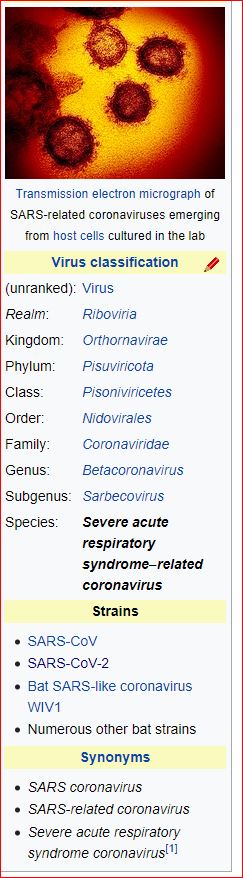
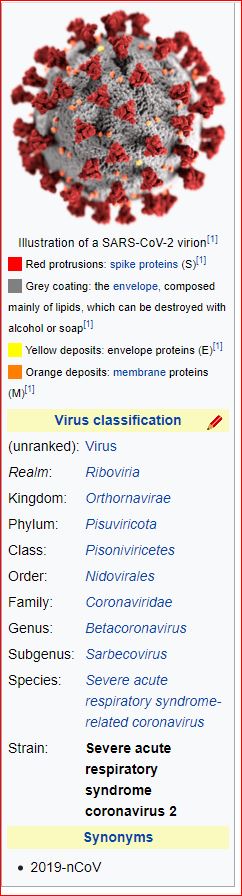
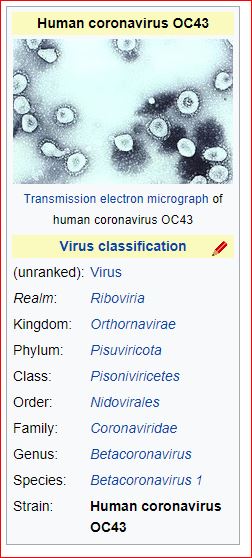
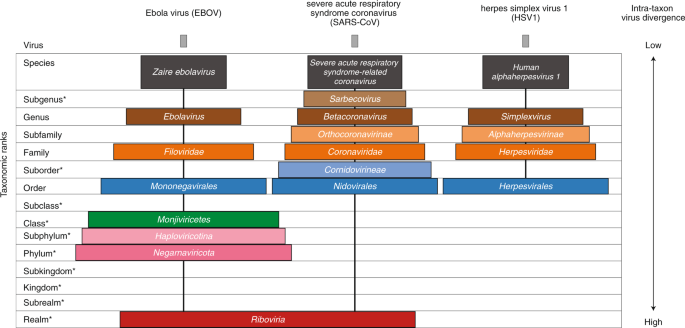
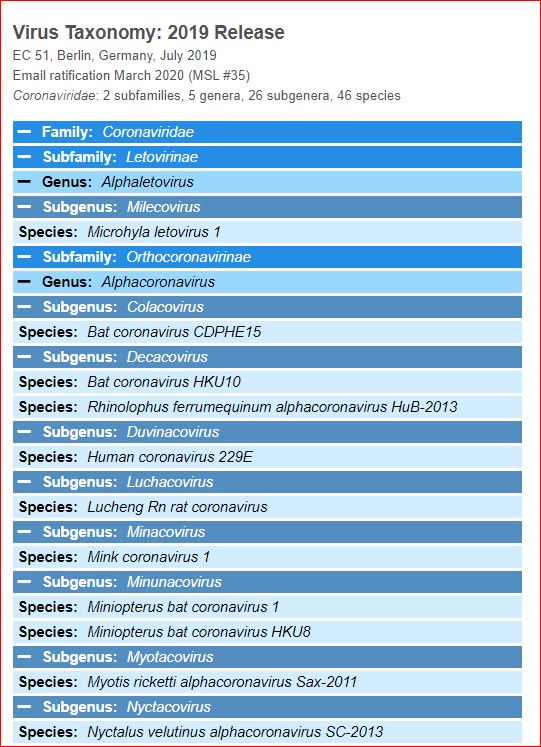
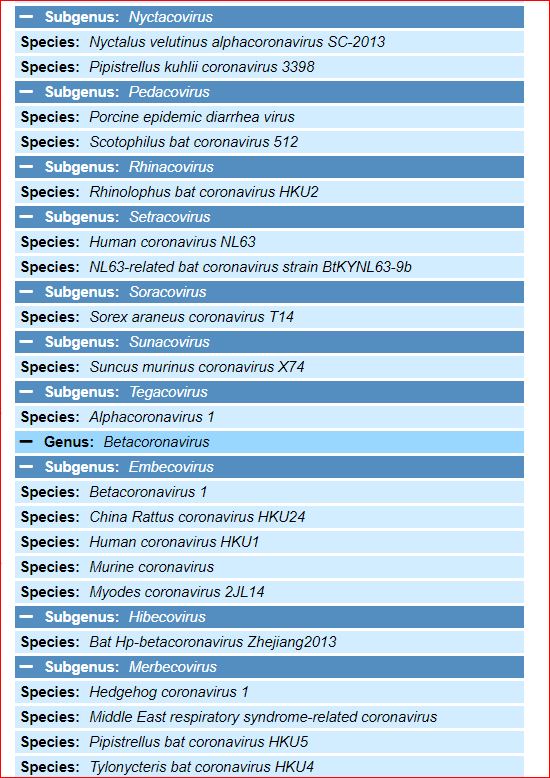
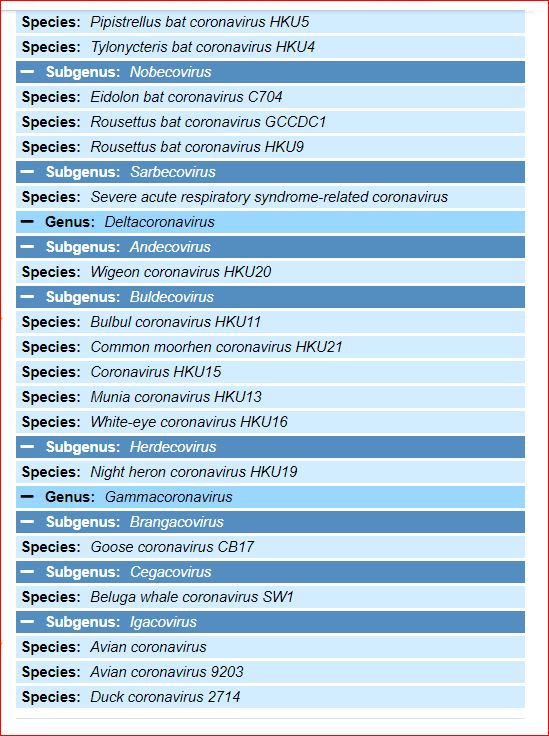
Application and impact of the extended virus taxonomy : When developing taxonomy, virologists are only obliged to assign a (new) virus to taxa at genus and species ranks. Other ranks may be used optionally when the scientific justification is sufficient. The new 15-rank taxonomic structure applies to all viruses, although none have been assigned to all ranks thus far. To illustrate the application of some of the newly established ranks, Fig. 2 and Table 1 show the full current classification and taxa demarcation criteria of two viruses with RNA genomes, Ebola virus (EBOV) and severe acute respiratory syndrome coronavirus (SARS-CoV). Both viruses are well-known human pathogens and members of the species Zaire ebolavirus and Severe acute respiratory syndrome-related coronavirus, respectively. Although both of these viruses infect humans, they differ taxonomically in the ranks that are populated and the demarcation criteria that define the taxa. Only at the basal, realm rank are the two viruses included in the same taxon, Riboviria. Figure 2 also shows the corresponding information for another human pathogen, herpes simplex virus 1, which has a double-stranded DNA genome and is currently assigned to taxa of five ‘traditional’ ranks. ( REFERERENCES, Aclnowledgements, contributors, etc )
Fig. 2: Classification of EBOV, SARS-CoV and herpes simplex virus 1 in the 15-rank taxonomic hierarchy.
Intra-cluster virus divergence, which increases from the virus to the species rank to the realm rank, is represented by the increasing width of the respective rectangles, which are not drawn to scale. EBOV is most closely related to, but distinct from, Bombali, Bundibugyo, Reston, Sudan and Taï Forest viruses, which belong to separate species included in the Ebolavirus genus. SARS-CoV is one of several closely related coronaviruses isolated from humans and animals, such as palm civets and bats, and are included in the species Severe acute respiratory syndrome-related coronavirus. Herpes simplex virus 1 is one of two human herpesviruses belonging to different species in the Simplexvirus genus. Ranks that were introduced with the extended rank structure are indicated by an asterisk.
A surge of activity to populate the existing and additional ranks of virus taxonomy can now be expected and is to be encouraged. This will include the consideration of numerous supergroups and superfamilies that have remained outside of virus taxonomy to date. At the time of writing (January 2020), the ranks included highly different numbers of taxa (Fig. 1). As expected, the species rank, with the lowest degree of intra-taxon virus divergence, is the most heavily populated, and ranks with higher degrees of intra-taxon divergence are the least populated. This highly uneven distribution of the number of taxa assigned at different ranks is due not only to the hierarchical relationship of the ranks, but also to the very recent introduction of the more basal ranks, and the lack of requirement to fill these ranks. The uneven distribution also reflects variations in the sampling of different virus lineages in diverse hosts as well as differences in the approaches adopted for the recognition of taxa in particular species by different ICTV Study Groups. ... These variations could persist, although the currently observed differences in taxon density may be partially alleviated when researchers define the more distant taxonomic relationships among viruses and improve their resolution by involving traditional and new evolutionary methodologies, such as network analyses28. For example, increasing evidence supports an ancestral relationship of some viruses of the order Caudovirales (a group of bacterial viruses with double-stranded DNA genomes) and viruses of the order Herpesvirales (a group of animal viruses with double-stranded DNA genomes) through a shared virion morphogenesis module. This module includes the HK97-type major capsid protein, portal protein, capsid maturation protease and the genome-packaging terminase complex25,39,40,41, and appears to reflect monophyletic relationships that may warrant taxonomic recognition. ... As a result of the change to the number and scope of ranks, virus taxonomy is now, for the first time, able to accommodate taxa at any level of virus divergence between the very narrow (species) and the extremely wide (realms). How these ranks are used will depend on the research community, including the ICTV. We stress that the validity of any established taxa, or those created in the future, depends on the strength of scientific evidence to support the demarcation and ranking of taxa, which is considered on a case-to-case basis. ... The codified availability of a greater number of ranks in a formal virus classification that emulates a Linnaean framework may also facilitate the comparison, and possibly improve the compatibility of virus taxonomy with the taxonomies of cellular organisms. Although the switching of hosts by viruses may be a complicating factor, the availability of fossils and a defined evolutionary timescale for some virus hosts should benefit virus taxonomy. Such information will be essential for taxa demarcation and rank definition in the future, notwithstanding that all taxonomies depend on the accuracy of evolutionary reconstructions, which are most challenging for distant relationships that reflect numerous changes, including those resulting from horizontal (lateral) gene transfer. ... We expect that the described changes to the hierarchical rank structure will create a new impetus for the exploration of virus macroevolution and a framework for its application to taxonomy. The changes will also stimulate research on the defining characteristics of monophyletic virus lineages and the recognition of historical events that played a decisive role in their origins and evolution. These events may be comparable to major transitions in the evolution of cellular life forms, such as the origins of eukaryotes or plants. This information could be used to define taxa and ranks, and the improved interaction with evolutionary research will facilitate the main mission of virus taxonomy, which is to serve the virology community and the public at large in a comprehensive, scientific manner.
Rights and permissions : Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
SOURCE: https://en.wikipedia.org/wiki/Viral_disease
"... Basic structural characteristics ...
- There are five double-stranded DNA families: three are non-enveloped (Adenoviridae, Papillomaviridae and Polyomaviridae) and two are enveloped (Herpesviridae and Poxviridae).
... All of the non-enveloped families have icosahedral capsids.
- There is one family of partly double-stranded DNA viruses: Hepadnaviridae. These viruses are enveloped.
-There is one family of single-stranded DNA viruses that infect humans: Parvoviridae. These viruses are non-enveloped.
- There are seven positive single-stranded RNA families: three non-enveloped (Astroviridae, Caliciviridae and Picornaviridae) and four enveloped (Coronoviridae, Flaviviridae, Retroviridae and Togaviridae).
... All the non-enveloped families have icosahedral nucleocapsids.
- There are six negative single-stranded RNA families: Arenaviridae, Bunyaviridae, Filoviridae, Orthomyxoviridae, Paramyxoviridae, and Rhabdoviridae. All are enveloped with helical nucleocapsids.
-There is one family with a double-stranded RNA genome: Reoviridae.
- There is one additional virus (Hepatitis D virus) that has not yet been assigned to a family but is clearly distinct from the other families infecting humans.
-There is one family and one genus of viruses known to infect humans that have not been associated with disease: the family Anelloviridae and the genus Dependovirus. Both of these taxa are non-enveloped single-stranded DNA viruses.
..."
 <
< 
{kind=link}